


Мы компания
производитель
Доставка по всей
России и СНГ
Наличная и
безналичная оплата
Существенные
оптовые скидки
Гарантия на
всю продукцию
Профессиональные
консультанты
Фитохром и криптохром при светодиодном led освещении для растений, гроубокса, теплиц
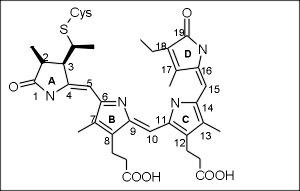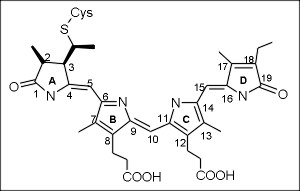
Физиологически важные области спектра, при освещении растений светодиодами. Фитохром и криптохром.
Поскольку растения заметно зависят от фотосинтеза, для регуляции процессов развития и дифференцировки они должны правильно оценивать количество и качество света. Заметим, что согласно физическим принципам есть предел энергии для преобразования в световых реакциях фотосинтеза, связанный обратно пропорциональной зависимостью с предельной длиной волны. Фотосистема I способна поглощать и преобразовывать кванты с длиной волны не более 700 нм, а фотосистема II - не более 680 нм. Если зеленые растения освещать квантами с длиной волны более, чем 700 нм, это приведет к гибели растения: в таких квантах недостаточно энергии для протекания реакций световой стадии фотосинтеза. Оптимальными для фотосинтеза будут длины волн в красной области в диапазоне 600-700 нм (они используются с наименьшими потерями). Таким образом, красные лучи можно разбить на две резко отличающиеся области - собственно красную, пригодную для фотосинтеза (до 700 нм) и дальнюю красную, для фотосинтеза не пригодную (более 700 нм). В принципе дальний красный свет и темнота должны приводить к одним и тем же реакциям - это сигнал того, что условий для успешного фотосинтеза нет, однако при дополнении данного спектра к основному при использовании светодиодных светильников для растений, позволяет добитьсялучших результатов. Для оценки качества и количества красной области падающего света у растений есть особый пигмент - фитохромобилин, аналогичный фикобилинам водорослей. Комплекс фитохромобилина с белком называют фитохромом.
Принцип работы фитохромной системы следующий. Фитохром может пребывать в двух основных состояниях, характеризуемых максимумом поглощения: Фкр.(660) и Фдкр.(730). Красная форма поглощает свет и превращается при этом в дальнюю красную форму. Переход Фкр. в Фдкр. под действием света с длиной волны 660 нм сигнализирует растению о том, что свет пригоден для фотосинтеза. Дальняя красная форма фитохрома может переходить в красную как при воздействии света с длиной волны 730 нм, так и самопроизвольно в темноте (самопроизвольный переход медленнее). Переход Фдкр. в Фкр - это сигнал о том, что условий для фотосинтеза нет (либо кванты света несут недостаточно энергии, либо света нет совсем). Таким образом, фитохромная система похожа на выключатель в электрической сети: если свет есть, то система ожидает сигнала о выключении, если света нет - система ожидает, чтобы его включили. В полной темноте чувствительность растений к свету обостряется: содержание определенных форм фитохрома заметно увеличивается. Как только этиолированные растения переносят на свет, содержание фитохрома быстро падает (но полностью фитохром не исчезает). Растение модулирует чувствительность к свету в зависимости от внешних условий. На основании сигналов, поступающих от фитохромной системы, растение принимает глобальные решения: готовиться к фотосинтезу или все запасы органических веществ пустить на рост, приступить ли к цветению или к листопаду, изменить ли стратегию роста, прорастать семенам или дождаться более благоприятных условий освещения и т.д. Вторая светочувствительная система связана с синей областью спектра. Там сосредоточены максимумы поглощения хлорофиллов, каротиноидов, флавинов и др. Таким образом, синяя область также физиологически важна. У растений синий свет воспринимают разнообразные рецепторы: криптохромы, фототропин. В отличие от красной части спектра, энергия синих квантов даже избыточна для фотосинтеза. Поэтому фоторецепторы синего света работают иначе: здесь нельзя выделить двух переключающихся состояний. Поступающие сигналы помогают принять локальные решения: определить направление максимальной освещенности при фототропизме, открыть устьица и т.д. В последнее время появляются данные о том, что у растений есть системы, чувствующие ультрафиолетовые лучи, однако механизмы ответов на ультрафиолет пока еще недостаточно изучены.
История открытия фитохрома.
В 1950-х годах американские исследователи Г.А.Бортвик, С.Б.Хендрикс и М.У.Паркер изучали прорастание семян салата 'Grand Rapids'. В полной темноте семена не прорастали даже если растениям были предоставлены влага и приемлемые температурные условия. Достаточно было лишь на несколько секунд осветить набухшие семена, и они прорастали. Набухшие семена салата были настолько чувствительными к свету, что приходилось работать с ними как с фотоматериалами, обертывая опытные образцы черной фотобумагой или светонепроницаемой фольгой. Бортвик и др. подробно получили спектр этой физиологической реакции, построив график зависимости % проросших семян от длины волны падающих квантов света. Кроме того, они обнаружили, что освещение дальним красным светом с длиной волны 730 нм обычно тормозит реакцию прорастания. Последовательное освещение красным (660) - дальним красным (730) - красным (660 нм) приводило к прорастанию семян. Но если последнюю вспышку света снова сделать дальней красной (730), то прорастание ингибировалось. Открытое явление получило название К/ДК- обратимости. Оно заключается в том, что ответ на низкую интенсивность освещения можно обратить: если действовал красный свет, ответ можно ингибировать дальним красным, и наоборот. Растения дают тот ответ, который можно вызвать последней вспышкой света. Была выдвинута гипотеза, что в семенах присутствует пигмент, со спектральными свойствами отличными от хлорофилла, который обладает свойством "переключаться" при воздействии внешнего стимула: красного или дальнего красного света. Гипотетический пигмент был назван фитохромом (phyton - растение, chromos - краска, пигмент). Спектр физиологической реакции прорастания семян позволил начать поиски и выделение пигментов, обладающих таким же спектром. Когда из растений выделяли очередной пигмент (или окрашенную фракцию), его спектр сравнивали с полученным физиологическим спектром и проверяли, годится ли пигмент на роль фитохрома.
Эта стратегия поиска увенчалась успехом: всего через 7 лет У.Л.Батлер (под руководством Хендрикса) выделил фитохром в чистом виде и охарактеризовал его. Фотоконверсия фитохрома. Фитохром А и фитохром В. Фитохром оказался димерным белком, содержащим ковалентно связанную хромофорную группировку из четырех пиррольных колец (А, В, С и D), соединенных метиновыми мостиками. Молекула хромофора синтезируется сначала как замкнутая тетрапиррольная структура (гем), а затем происходит размыкание и встраивание в белок. Как и ожидалось, хромофорная группа фитохрома (фитохромобилин) существует в двух основных спектральных формах, которые могли переходить друг в друга под действием квантов света. Форма Ф660 (красная) улавливала красные кванты света и переходила в Ф730 (дальнюю красную). Это объясняло эффект К/ДК-обратимости: при освещении красным светом форма Ф660 исчезала (форма в Ф730 накапливалась). Если после этого дать свет с длиной волны 730, форма Ф730 исчезнет (перейдет в Ф660). Последовательные вспышки приведут к тому, что сначала весь фитохром переходит в одну форму, а потом в другую и т. д. Для физиологической реакции важна именно последняя форма фитохрома, т. е. последняя короткая вспышка, которой освещали растение. Дальнейшие исследования показали, что главным событием, приводящим к превращению форм фмтохрома, является поворот кольца D относительно кольца C (цис-транс изомеризация в метиновом мостике, соединяющем эти кольца). Эта первичная реакция приводит к деформации белкового кармана и изменению структуры димера вцелом. Первичная форма возбужденной хромофорной группировки оказывается недостаточно стабильной и самопроизвольно через ряд этапов переходит в более стабильные формы, причем промежуточные формы для перехода из Ф660 в Ф730 отличаются от промежуточных форм для обратного перехода. В 1990-х годах были изучены гены, отвечающие за биосинтез белкового компонента фитохрома. Оказалось, что у большинства объектов преобладающими ялвяются фитохром А и фитохром В, отличающиеся не только свойствами белковой части, но и спектром поглощения (эффект различного белкового окружения хромофора). Кроме двух преобладающих фитохромов, показано, что в растениях имеются минорные формы: фитохром С, фитохром D и фитохром E. Роль минорных форм фитохрома в регуляции ответа на свет еще недостаточно изучена.
При всем различии белковой части фитохромов, в белковой молекуле можно выделить высококонсервативный фитохромобилин-связывающий домен, расположенный ближе к N-концу белка. При повреждении N-концевого участка хромофор не может встроиться в молекулу фитохрома. С-конец содержит участки, похожие на бактериальные гистидин-киназы (обычно имеется два гистидин-киназных домена). По-видимому, фитохром обладает киназной активностью, которая зависит от состояния хромофорной группировки. Кроме того, фитохром может фосфорилироваться, меняя свою активность. Более того, предполагают, что фитохром может фосфорилировать сам себя. Фитохром А обладает более широкими пиками поглощения, а фитохром В - более узкими. Так, например, в спектре Ф(А)660 несмотря на то, что максимум поглощения равен 660, широкое плечо выходит и в дальнюю красную область спектра. С точки зрения рецепции света это означает, что фитохром А можно перевести в форму Ф(А)730 не только облучением квантами с длиной волны 660, но и квантами из дальней красной области (730). Правда, ДК-квантов для этого перехода нужно намного больше, чем квантов, соответствующих максимуму поглощения. Следовательно, растение должно воспринимает одинаково как низкие интенсивности красного света (660), так и высокие интенсивности дальнего красного (730). Фитохром А отвечает за ответы на ДК-свет высокой интенсивности (HIR). В спектре поглощения фитохрома В пики более узкие, у формы Ф(В)660 плечо не заходит в дальнюю красную область. Это означает, что Ф(В)660 нельзя перевести в форму Ф(В)730 с помощью интенсивного дальнего красного света. Фитохром В отвечает за ответы на низкую интенсивность и обеспечивает при этом эффект К/ДК-обратимости. Наличие двух фитохромов, заметно отличающихся по спектральным характеристикам, позволяет растению различать (1) полную темноту, (2) интенсивный ДК-свет и (3) хорошее освещение, пригодное для фотосинтеза. У растений, выращенных в полной темноте, повышено содержание фитохрома А - именно этот фитохром отвечает за восприятие света любого качества - растение "ищет" свет, и "старается" повысить чувствительность к нему. При переносе на свет (независимо от того, К или ДК), концентрация фитохрома А начинает быстро снижаться до уровня, характерного для выросших на свету растений. (У этиолированных растений иногда выделяют пул фитохрома А', который будет разрушен при получении светового сигнала достаточной интенсивности, и собственно пул фитохрома А, который сохраняется на свету). Самыми ранними событиями, которые удается зарегистрировать при воздействии на клетки красным или дальним красным светом, являются К/ДК-обратимые изменения мембранного потенциала. Эти явления были впервые обнаружены на зеленых водорослях и часто сопровождаются быстрым изменением положения хроматофора.
В качестве возможных вторичных мессенджеров фитохромного сигнала рассматривают G-белки, которые формируют гетеротримеры, циклическую ГМФ и кальций. Эксперименты с растениями, у которых ген фитохрома соединен с геном зеленого флуоресцирующего белка (Green Fluorescent Protein) показали, что в зависимости от освещения фитохром может находиться либо в цитоплазме, либо в ядре. Как правило, если растения выращены в темноте, то фитохром находится в цитоплазме. Скорость поступления и световые стимулы, необходимые для перемещения фитохрома А и фитохрома В в ядро несколько отличаются. Так, для перемещения фитохрома В в ядро необходим красный свет. Это означает, что в конформации Ф(В)650 не может пересекать ядерную мембрану, а после фотоконверсии в форму Ф(В)730 в течение примерно 30 минут аккумулируется в ядре. Фитохром А переходит в ядро как при освещении как красным, так и дальним красным светом высокой интенсивности. Интересно, что на С-концевом участке у фитохромов есть два PAS-домена, которые встречаются в транскрипционных факторах. PAS-домены важны как для ДНК-белковых, так и для белок-белковых взаимодействий. Повреждение хотя бы одного из PAS-доменов приводит к нарушению функционирования фитохрома. Это означает, что проникая в ядро, фитохром может непосредственно участвовать в регуляции транскрипции, связываясь в комплексы с различными транскрипционными факторами.

")

")